|
ИНСТИТУТ ПРИКЛАДНОЙ МАТЕМАТИКИ им. М.В. КЕЛДЫША РОССИЙСКОЙ АКАДЕМИИ НАУК
На правах рукописи
БУРЦЕВ Михаил Сергеевич
ИССЛЕДОВАНИЕ НОВЫХ ТИПОВ САМООРГАНИЗАЦИИ И ВОЗНИКНОВЕНИЯ ПОВЕДЕНЧЕСКИХ СТРАТЕГИЙ
05.13.18 – математическое моделирование, численные методы и комплексы программ
Автореферат диссертации на соискание ученой степени кандидата физико‑математических наук
Москва – 2005
Работа выполнена в Институте прикладной математики им. М.В. Келдыша Российской академии наук
Научные руководители: доктор физ.‑мат. наук Редько Владимир Георгиевич доктор физ.‑мат. наук, профессор Малинецкий Георгий Геннадьевич Официальные оппоненты: доктор физ.‑мат. наук, академик РАН Бугаев Александр Степанович доктор физ.‑мат. наук, профессор Смолянинов Владимир Владимирович
Ведущая организация: Институт системного программирования РАН
Защита диссертации состоится «28» июня 2005 г. на заседании диссертационного совета Д002.024.02 при Институте прикладной математики им. М.В. Келдыша РАН по адресу: Москва, Миусская пл., д. 4.
С диссертацией можно ознакомиться в библиотеке Института прикладной математики им. М.В. Келдыша РАН
Автореферат разослан «17» мая 2005 г.
Ученый секретарь диссертационного совета
Г.В. Устюгова I. Общая характеристика работыАктуальность темы. Динамика многих биологических и социальных систем определяется эволюцией в популяциях, поведение агентов в которых сильно варьируется. Широкий спектр стратегий поведения и типов взаимодействий агентов делает невозможным применение для моделирования подобных систем стандартных методов статистики и теории игр. Эта проблема привела к появлению отдельного класса компьютерных моделей, получившего название – "Искусственная жизнь". Главными отличиями моделей искусственной жизни является то, что в них агент обладает широким спектром возможных стратегий поведения, и его приспособленность не задана явно, а определяется в процессе "жизни" – взаимодействия со средой и другими агентами. Такой подход позволяет отказаться от субъективного понимания исследователем меры приспособленности агента (что "хорошо", и что "плохо" для выживания агента), и перейти к анализу эволюционно устойчивых состояний системы популяция-среда. При этом модель может обладать поведением, по сложности приближающимся к прототипу, но в отличие от экспериментов с реальной живой или социальной системой (которые не всегда осуществимы), эксперименты с компьютерной моделью легко воспроизводимы, и все изменения в эволюции системы могут быть зафиксированы для последующего анализа. Таким образом, модели искусственной жизни могут быть использованы непосредственно при исследовании сложных систем, или в качестве полигона для исследования аналитических моделей. Исследования в рамках искусственной жизни являются продолжением пионерских работ по теории самовоспроизводящихся автоматов [1], игр автоматов [2] и компьютерного моделирования эволюции [3]. С момента своего возникновения, искусственная жизнь тесно связана с синергетикой и нелинейной динамикой, не случайно первый сборник статей в этой области вышел в журнале Phisyca D "Nonlinear Phenomena" [4]. И сегодня, синергетика и искусственная жизнь, широко использующая представления о самоорганизации и динамических системах, имеют общее направление движения [5,6]. Методы искусственной жизни активно используются в работах по моделированию эволюции когнитивных способностей животных [7], в областях эволюционной роботики и эволюционных вычислений [8]. Применение искусственной жизни позволяет подвергнуть теоретические эволюционные концепции более жесткому тесту. Так, в исследованиях эволюции кооперации, базирующихся на наиболее распространенном сегодня подходе теории игр, репертуар возможных взаимодействий между агентами очень беден, обычно агент может выбрать одно из двух действий – кооперироваться или не кооперироваться с текущим партнером. Искусственная жизнь позволяет смоделировать эволюцию популяции с гораздо более широкими и естественными возможностями для взаимодействия агентов, что дает гораздо более широкий спектр стратегий поведения агентов. Учитывая, что сегодня в области искусственной жизни не существует моделей эволюции родственной кооперации, в качестве задачи для моделирования в данной работе была выбрана проблема эволюции кооперации основанной на наследуемых маркерах. На сегодня серьезной проблемой в области искусственной жизни является то, что желаемый объем опыта по прикладному моделированию и интерпретации результатов вычислительных экспериментов еще не накоплен. Для того чтобы исследовать возможность использования искусственной жизни для моделирования реальных социальных систем, в работе была предпринята попытка построить модель для предсказания основных характеристик и динамики эволюции в примитивных эгалитарных сообществах охотников-собирателей и ранних земледельцев. Важной задачей раздела биологии, занимающегося адаптивным поведением, является понимание механизмов эволюционного возникновения целенаправленного поведения и его эволюционной устойчивости. Возникновение целенаправленности можно рассматривать, как один из важнейших этапов эволюционного развития когнитивных способностей организмов [7]. Метод искусственной жизни является одним из наиболее адекватных инструментов для исследования этой проблемы благодаря своей ориентированности на эволюцию и адаптивность поведения отдельных агентов. Это обусловило выбор проблемы эволюции целенаправленного поведения в качестве объекта для моделирования в данной работе. Цели работы. 1. Построение и исследование эволюционной компьютерной модели возникновения кооперации агентов. 2. Моделирование социальной эволюции в примитивных сообществах охотников-собирателей и ранних земледельцев. 3. Разработка компьютерной модели эволюции целенаправленного поведения. 4. Разработка и формальное описание некоторого класса и конкретной модели искусственной жизни. 5. Разработка методов измерения и визуализации динамики эволюционных компьютерных моделей. Методы исследования. Основным методом исследования является построение моделей искусственной жизни и вычислительный эксперимент с этими моделями. В работе было предложено четыре варианта эволюционной модели кооперации: без агрессии и наследуемых маркеров; с агрессией и без маркеров; с агрессией и маркерами; аналог предыдущего варианта, но с высоким уровнем летальности макромутаций, а так же модель целенаправленного поведения. Для анализа результатов расчетов модели использовались предложенные в работе методы обработки и визуализации данных, направленные на поиск и исследование траектории эволюционирующей системы в пространствах генотипов и стратегий поведения. Для интерпретации результатов вычислительных экспериментов с моделью кооперации использовалась база данных, составленная по результатам антропологических и этнографических исследований. Научная новизна. Впервые в классе моделей искусственной жизни предложена и исследована модель эволюции кооперации, базирующейся на наследуемых маркерах. Исследование результатов компьютерного моделирования привело к выявлению новых стратегий поведения неизвестных в эволюционной теории игр. Впервые для данного класса моделей проведено моделирование социальной динамики с последующей верификацией результатов численных экспериментов на соответствие данным кросс-культурных исследований. Предложена и исследована оригинальная модель эволюции целенаправленного поведения основанного на мотивациях. На основе формализма теории автоматов разработан новый способ описания конкретных моделей искусственной жизни и их классов. В работе предложен новый метод исследования динамики моделей искусственной жизни, базирующийся на теории динамических систем. Эволюционирующая популяция агентов рассматривается как динамическая система, для которой ищутся и анализируются траектории в различных фазовых пространствах. Практическая ценность. Полученные результаты используются в Институте прикладной математики им. М.В. Келдыша РАН, Институте оптико-нейронных технологий РАН и Центре цивилизационных и региональных исследований РАН. Построенные модели могут быть использованы при исследовании эволюции социальных систем, в которых кооперативные отношения играют важную роль. Разработанные технологии являются основой для построения программных многоагентных систем, предназначенных для распределенной интеллектуальной обработки информации. Результаты работы могут использоваться при разработке подобных систем и повысить их эффективность. Апробация работы. Основные результаты работы докладывались на конференциях "Биосфера и человечество" (Обнинск, 2000), 8‑й Международной конференции "Математика. Компьютер. Образование" (Пущино, 2001), 3-й, 4-й и 7-й Научно-технической конференции "Нейроинформатика" (Москва, 2001, 2002, 2005), Международной конференций "Искусственные интеллектуальные системы" (IEEE AIS, Геленджик, 2002), 8-ой Национальной конференции по искусственному интеллекту с международным участием (Коломна, 2002), 4-й Международной конференции "European conference on Artificial Life" (Дортмунд, 2003), Международной конференции "Математическое моделирование социальной и экономической динамики" (Москва, 2004), 3-ей Международной конференции "Иерархия и власть в истории цивилизаций" (Москва, 2004), на международных школах "Summer University Complex Systems course" (Будапешт, 2001) и "Complexity in Social Sciences" (Баеза, 2003), на семинарах Института проблем передачи информации РАН, Института системного программирования РАН, Института оптико-нейронных технологий РАН, Института нормальной физиологии им. П.К. Анохина РАМН, Института высшей нервной деятельности РАН, Центрального экономико-математического института РАН, Института прикладной математики им. М.В. Келдыша РАН. Публикации. По результатам выполненной работы имеется 26 публикации (см. список публикаций). Структура диссертации. Диссертация состоит из введения и четырех глав, изложенных на 120 страницах, содержит 29 рисунков, 14 таблиц и библиографию из 181 наименования. II. Содержание работыВо введении излагаются цели работы. Показывается место искусственной жизни среди других методов моделирования. Дается обзор существующих подходов к моделированию эволюции биологических и социальных систем. Ставятся задачи и обосновываются методы моделирования. Приводится краткое изложение содержания диссертации. Первая глава посвящена формализации некоторого класса моделей искусственной жизни и методам их исследования. В первой части главы рассматриваются существующие математические и компьютерные модели в области эволюционной биологии. Во второй представлено изложение оригинальной формализации некоторого класса моделей искусственной жизни и методы их исследования. Модели искусственной жизни концептуально близки к моделям коллективного поведения автоматов, что позволяет позаимствовать стиль описания из теории автоматов [2]. Рассмотрим популяцию P агентов A, эволюционирующую в среде U. Поведение отдельного агента A задается следующим образом:
здесь j – вектор внутренних состояний агента A; s – вектор входных переменных; f – выходная переменная. В уравнениях (1) и (2) переменная t имеет смысл времени и принимает целочисленные значения t = 1, 2, . . . Значения входных переменных s определяются текущим состоянием, окружающей агента среды. Будем считать, что выходная переменная f принимает n дискретных значений f1, f2, . . . , fn, и называть эти значения действиями. Так если в момент времени t выходная переменная принимает некоторое значение f(t) = fk мы будем говорить, что в момент t агент A произвел действие fk. Определим зависимость внутреннего состояния ji(t) агента A от входных переменных s(t) как взвешенную сумму, тогда выражение (1) принимает вид: j(t) = Ws(t), (3) где W – матрица весов, компоненты которой могут принимать значения из некоторого ограниченного интервала [-Wmax;Wmax]. Для удобства в работе рассматриваются агенты, у которых число внутренних состояний ji равно числу возможных действий n, что позволяет определить (2) следующим образом: f(t) = fk , Последовательное применение (3) и (4) аналогично функционированию простой искусственной нейронной сети. Будем считать, что каждый агент A популяции P обладает ресурсом r. Изменение значения ресурса агента зависит от его действий:
Коэффициент ki имеет отрицательное значение, если действие приводит к уменьшению ресурса агента, и положительное в противном случае. Для каждого момента времени
t после вычисления
действий и изменений значений ресурсов для всех агентов популяции P, отбираются агенты, значение
ресурса у которых меньше или равно нулю Для обеспечения воспроизводства популяции в репертуаре действий агента имеется действие "делиться" fd. Если агент Ap (родитель) совершает действие fd, то в популяцию P добавляется новый агент Ao (потомок). Матрица весов потомка Wo формируется на основе матрицы весов родителя Wp следующим образом: Woi,j = Wpi,j + ξi,j, (6) где ξi,j – случайная величина, имеющая равномерное распределение на интервале [-pξ;pξ]. Выражение (6) задает мутационный процесс с амплитудой мутаций pξ. Будем называть матрицу W – генотипом агента A, отдельный элемент матрицы Wij – геном. Стратегией поведения агента A или его фенотипом будем называть отображение из пространства входных в пространство выходных переменных (действий), задаваемое (1) и (2). Взаимодействие между агентами определяется топологией среды U. Основу U составляет ограниченное множество дискретных клеток, в которых могут находиться агенты A. Топология U накладывает ограничения на возможные переходы агентов между клетками. Для того чтобы смоделировать эволюцию популяции на плоскости, в предложенной модели клетки среды были помещены в узлы двумерной прямоугольной сетки, натянутой на тор. Таким образом, за один такт времени агент мог переместиться в одну из четырех соседних клеток среды U. Предложенная формализация облегчает описание и анализ конкретных моделей искусственной жизни, и была использована во второй и четвертой главе диссертации. В заключительном разделе первой главы дается краткий обзор методов анализа моделей искусственной жизни, а затем приводится изложение оригинального подхода, основывающегося на представлении популяции, как динамической системы. Предложенный подход в дальнейшем используется в работе для анализа результатов компьютерных экспериментов с моделью кооперации. Во второй главе предложена модель искусственной жизни, предназначенная для исследования эволюции кооперации, приведены и проанализированы результаты моделирования. Кооперативное поведение животных хорошо вписывается в эволюционную теорию, если кооперирующиеся особи принадлежат разным видам, и взаимоотношения между ними носят характер симбиоза. Но кооперация также широко распространена и между особями одного вида, несмотря на то, что они занимают одну и ту же экологическую нишу, и, следовательно, конкуренция между ними максимальна. Этот феномен ставит серьезную проблему перед теорией естественного отбора. Решение, объясняющее возможность возникновения кооперации между генетически родственными организмами, было предложено В. Гамильтоном [9]. Оно заключается в том, что кооперация возможна, если выполнено условие:
где r – мера генетической близости, b – выигрыш объекта кооперативного поведения, а c – затраты самого животного. Эволюционные механизмы, приводящие к кооперативному поведению между родственниками, получили название родственного отбора. При помощи аппарата теории игр было показано, что для ряда представляющих интерес ситуаций, кооперация между родственниками будет эволюционно устойчивой стратегией [10]. Обычно теория игр работает с моделями с малым количеством исходов в игре, а, следовательно, с простой структурой выигрышей, кроме того, часто предполагается равномерное перемешивание игроков, что не позволяет учесть локальные пространственные взаимодействия, являющиеся важным свойством многих реальных систем. Поэтому представляет интерес исследование родственного отбора при помощи модели искусственной жизни, в которой агенты обладают достаточно большим набором стратегий поведения и взаимодействуют в пространстве. В предложенной модели популяция P агентов A помещалась в клеточную среду, в клетках которой с некоторой вероятностью появлялся ресурс, необходимый агентам для совершения действий. Для того чтобы агенты могли отличать родственников, каждому агенту A был приписан вектор-маркер m, значения которого наследовались с мутациями от агента-родителя Ap. Компоненты вектора m принимали целочисленные значения из некоторого интервала [-Mmax;Mmax]. Вектор s значений входных переменных агента формировался на основе информации о локальном окружении, значении внутреннего ресурса r и евклидовом расстоянии между собственным маркером m и маркером соседа по клетке. Значения выходной переменной f, определяющей действия агента и соответствующих коэффициентов ki изменения внутреннего ресурса агента, использовавшиеся при моделировании, приведены в таблице 1. Эволюционный алгоритм, изложенный в первой главе, был несколько модифицирован. В геном g агента A, содержащий матрицу весов W, были добавлены два бинарных вектора I и O, кодировавших наличие компонент в векторе входных переменных s и значений выходной переменной f. Компоненты векторов I и O подвергались мутациям малой величины при делении агента Ap, таким образом, существовала малая вероятность, что потомок Ao будет иметь набор входных переменных и действий несколько отличный от соответствующего набора родителя. Мутации векторов в модели интерпретировались, как аналог биологических макромутаций. Изменения значений компонент векторов I и O приводили к тому, что при формировании матрицы весов Wo потомка Ao в матрицу родителя Wp добавлялись или удалялись столбцы или строки соответственно. Таблица 1
* при моделировании коэффициенты ki были заданы в соответствии со следующими равенствами: k2 = 2k1, k4 = k5 = 2k2, k6 = 25k4, коэффициент k3 являлся параметром в серии экспериментов. При делении (f = f5) после изменения энергии родителя (Dr = -k5) половина его ресурса передается потомку. Компьютерные эксперименты проводились с четырьмя вариантами модели (см. таблицу 2). Таблица 2
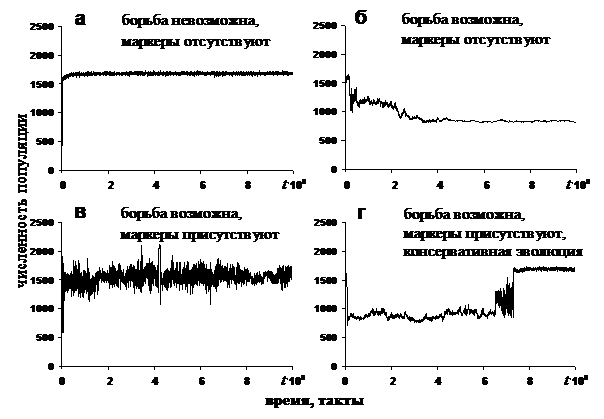
В каждом эксперименте начальная популяция P состояла из агентов A, которые имели одинаковые геномы g{I;O;W}. Геном агента начальной популяции был задан таким образом, что агент мог получать информацию, о наличии ресурса в текущей клетке, клетке по направлению "вперед", и совершать только три действия: потреблять ресурс, двигаться вперед и делиться. С моделью была проведена серия экспериментов. Для каждого из вариантов модели (см. табл. 2), при различных значениях эффективности потребления ресурса k3, производился расчет трех версий, отличавшихся начальным значением генератора случайных чисел, все остальные параметры были зафиксированы. В каждом эксперименте эволюция длилась 5×107 тактов, что значительно больше характерного времени перехода модели в стационарный режим с постоянной численностью популяции или в динамический режим с регулярными колебаниями численности. На рисунке 1 представлены результаты компьютерных экспериментов. На графике отображены установившиеся значения численности популяции NSi для всех вариантов модели (i = 1,2,3,4) в зависимости от эффективности потребления ресурса k3. Для первого варианта модели (см. табл. 2), в котором агенты не могут отнимать ресурс друг у друга, точки ложатся на прямую. Это значит, что максимальная численность популяции растет прямо пропорционально эффективности потребления ресурса NS1 = NSmax µ k3. Типичная динамика численности популяции для первого варианта модели приведена на рис. 2а. Зависимость NS2 (k3), полученная в экспериментах с вариантом модели, в котором агенты могут нападать друг на друга, но не могут отличать родственников (вариант 2 из табл. 2), распадается на два участка (рис. 1). Для тех значений k3, для которых максимальная численность популяции меньше числа клеток в среде NC (NC = 900 во всех экспериментах) NSmax(k3) ≤ NC, т.е. одна клетка не способна "прокормить" одного агента, численность популяции NS2 (k3) = NSmax(k3) = NS1 (k3). Для значений k3, для которых выполняется условие NSmax(k3) > NC, численность популяции NS2 (k3) = NС. Пример динамики численности популяции для последнего случая (NSmax(k3) > NC) приведен на рис. 2б. Независимость установившейся численности NS2 от k3 для NSmax(k3) > NC вызвана тем, что в популяции доминирует стратегия поведения, в соответствии с которой два агента, находящиеся в одной клетке, нападают друг на друга и продолжают бороться, пока один из них не погибнет.
Рис. 1. Зависимость установившегося значения численности популяции NSi от эффективности потребления ресурса k3 для различных вариантов модели (i = 1,2,3,4). Поведение третьего варианта модели (см. табл. 2), в котором агенты могут отнимать друг у друга ресурс и имеют маркеры, позволяющие отличать родственников, при значениях k3, для которых выполняется условие NSmax(k3) ≤ 2NC, совпадает с поведением второго варианта модели NS3 (k3) = NS2 (k3). При значениях k3, для которых NSmax(k3) ³ 2NC численность популяции постоянно колеблется. Типичное поведение численности популяции во времени для этого случая приведено на рис. 2в. Кроме того, при увеличении k3, максимальная амплитуда колебаний численности растет (на рис. 1 интервалы колебаний численности для точек NS3 показаны вертикальными пунктирными линиями). Причины, вызывающие колебания, будут рассмотрены ниже. Компьютерные эксперименты с четвертым вариантом модели (см. табл. 2) показали, что NS4 (k3) = NS3 (k3) = NS2 (k3) для значений k3 удовлетворяющих условию NSmax(k3) £ 2NC, и NS4 (k3) = NS1 (k3) = NSmax (k3) для значений k3 удовлетворяющих условию NSmax(k3) ³ 2NC. Наиболее интересное поведение динамики численности популяции для этого варианта модели наблюдается при значениях k3, для которых NSmax(k3) » 2NC. В этом случае динамика численности популяции сначала соответствует второму варианту модели, а затем "переключается" в режим соответствующий первому варианту модели. Пример такого "переключения" представлен на рис. 2г.
Рис. 2. Динамика численности популяции для различных вариантов модели (см. таблицу 2). Для случаев а), б), г) эффективность потребления ресурса k3 = 1000, для случая в) k3 = 1250. Для того чтобы понять, как родственный отбор влияет на эволюцию в модели, рассмотрим подробнее эксперимент с четвертым вариантом модели для значения эффективности потребления ресурса k3, удовлетворяющего условию NSmax(k3) » 2NC. Динамика численности популяции для этого эксперимента приведена на рис. 2г. Воспользуемся предложенным во второй главе диссертации методом исследования эволюции стратегий поведения в популяции. Родственность играет роль при взаимодействиях агентов, поэтому будем рассматривать поведения агентов только в тех ситуациях, когда в клетке находятся два агента, которые могут быть родственниками (малое значение расстояния между маркерами m) или чужаками (большое значение расстояния между маркерами m). Кроме того, проследим, как будет меняться поведение агентов в этих ситуациях для разных значений накопленного ресурса r. Для этого зададим значения соответствующих компонент входного вектора s, и рассчитаем действия для каждого из агентов популяции P. Агентов, действия которых для всех заданных ситуаций попадают в одни и те же группы действий a (см. табл.3), будем считать придерживающимися одной и той же стратегии поведения. Стратегию поведения будем кодировать шестью разрядами, каждый из которых может принимать одно из четырех значений в соответствии с таблицей 3. Таблица 3
где a{0:"отдыхать,поворачиваться"; 1:"убегать"; 2:"нападать"; 3:"делиться"}, i = 1,2,3,4.
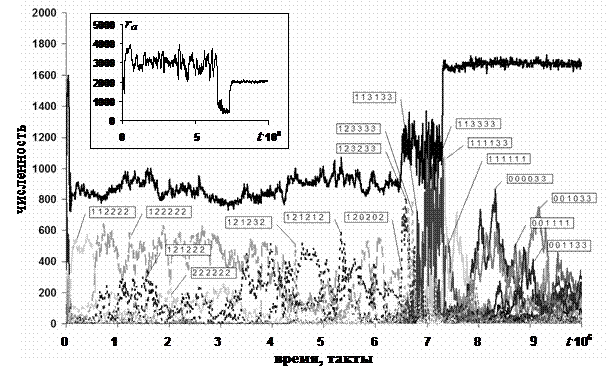
Рис. 3. Динамика численности агентов популяции, принадлежащих к различным стратегиям поведения (для определения стратегий поведения, закодированных на выносках, см. текст и табл. 3). На врезке показано изменение среднего по популяции значения внутреннего ресурса агентов ra от времени. Для удобства анализа разобьем всю эволюцию системы (рис. 3) на три периода: 1-ый с 0 по 6,5.106 такт, 2-ой с 6,5.106 по 7,5.106 такт, 3-ий с 7,5.106 по 107 такт времени. В течение первого периода численность популяции примерно равна числу клеток в среде, все агенты в популяции обладают той или иной агрессивной стратегией. Подавляющее большинство агентов следует стратегиям, заключающимся в избегании конкуренции с родственниками за ресурс и нападении на чужаков (стратегии типа "121212" ‑ покинуть клетку, если в ней родственник, и напасть, если чужак). Второй период начинается с появления в популяции новой кооперативной стратегии "120202". Следующие этой стратегии агенты, остаются в одной клетке с родственниками до тех пор, пока им хватает ресурса. Успешность такой стратегии связана с тем, что каждый агент в отдельности может владеть ресурсом r ≤ rmax, а, находясь в одной клетке, родственники в сумме могут накопить ресурс больший rmax, и, следовательно, при борьбе чужака с несколькими родственниками, вероятность выживания одного из родственников будет выше вероятности выживания чужака. Таким образом, если в первом периоде родственники кооперировались за счет избегания конкуренции за ресурс, то теперь они начинают кооперироваться за счет совместной обороны. Для совместной обороны выгодно иметь как можно больше родственников, что приводит к появлению и распространению быстро делящихся агентов (стратегии "123233" и "123333"), так как при делении половина ресурса родителя передается потомку, то средний по популяции ресурс быстро падает (см. врезку на рис. 3). В популяции начинают доминировать стратегии, когда агент не нападает на чужака, а пытается от него убежать (например, "113133"), при этом за счет высокой скорости деления ресурс таких агентов низок и нападение на них не выгодно (r ≤ k6). Все это приводит к вымиранию агентов с агрессивными стратегиями к концу второго периода. В начале третьего периода численность популяции быстро достигает максимального для данного k3 значения NSmax(k3). В третьем периоде агенты с агрессивными стратегиями отсутствуют, при этом в популяции присутствуют, как агенты с родственными стратегиями (например, "001033"), так и агенты, не отличающие родственников ("000033"). Для того чтобы облегчить анализ поведения модели выделим несколько крупных классов стратегий. При использовании теории игр для моделирования взаимодействия животных принято давать названия стратегиям поведения. Так, например, агенты, всегда проявляющие агрессивность в отношении партнера, называются «ястребы», а всегда избегающие конфликта – «голуби». Кроме ястребиных и голубиных стратегий в исследуемой модели присутствует стратегия «буржуа». Эта стратегия заключается в том, что агент не посягает на «собственность» других агентов, т.е. остается в своей клетке, но защищает «свой» ресурс, нападая на любого, пришедшего на его территорию. Наличие в модели возможности изменения поведения в зависимости от значения маркера партнера позволяет ввести пару новых стратегий[1]. В одну из них попадают агенты, избегающие конкуренции за ресурс с родственниками (малое расстояние между маркерами) и нападающие на чужаков. Примером такой стратегии может служить стратегия, закодированная в соответствии с таблицей 3 как «121212». В соответствии с поговоркой: «Ворон ворону глаз не выклюет», будем называть эту стратегию стратегией «ворона». В природе у многих малых стайных птиц, например у скворцов, наблюдается кооперативная защита от хищника. Если провести аналогию с коллективной защитой в модели, то можно назвать агентов, придерживающихся подобной стратегии – «скворцами». Определения стратегий для исследуемой модели приведено в таблице 4. Таблица 4
Рассмотрим, как изменение эффективности потребления ресурса k3 влияет на эволюцию кооперации в третьем варианте модели (см. таблицу 2). Это вариант обладает наиболее интересной динамикой, кроме того, четвертый вариант модели можно рассматривать, как частный случай третьего варианта. При низких значениях эффективности потребления ресурса k3 < k3С (см. рис. 1) ресурса, появляющегося в одной клетке, недостаточно для выживания одного агента, поэтому агенты вынуждены постоянно перемещаться в пространстве. Постоянное движение агентов приводит к тому, что конкуренция за ресурс между родственниками отсутствует, а, следовательно, отсутствует и вызываемое ей давление отбора. Но если агенты могут нападать друг на друга, то присутствует родственный отбор в сторону не нападения на родственников. Поэтому при k3 < k3С в популяции частота агентов, способных дифференцировать свое поведение в отношении родственников, растет вместе с частотой агрессивных агентов (см. рис. 4а). Анализ стратегий показывает, что основная часть агрессивных агентов в этом интервале значений k3 являются воронами (см. рис. 4б). Когда одна клетка способна «прокормить» одного агента, но не двух, т.е. k3С < k3 < 2k3С, отдельный агент имеет тем большую вероятность выжить в данный момент времени, чем выше его внутренний ресурс, так как от величины накопленного ресурса зависит успешность борьбы. Это приводит к тому, что отбор действует в сторону повышения внутреннего ресурса, и отражается в приближении максимума распределения агентов по накопленной энергии к максимально возможным значениям. Так как все агенты обладают примерно одинаковым внутренним ресурсом, а значит и одинаковыми шансами на выживание в поединке, то в популяции устанавливается «баланс сил» – доминирующей стратегией является стратегия буржуа. При этом в популяции присутствуют, как агенты, не различающие родственников, так и вороны. В модели при делении агента энергия распределяется поровну между родителем и потомком, что делает каждого из них неконкурентоспособным в сравнении с агентами из соседних клеток. Следовательно, если один из агентов после деления покидает клетку, он с большой вероятностью погибает. С другой стороны, если они оба остаются в клетке, то даже в случае ненападения друг на друга один из них погибает от недостатка ресурса (т.к. k3 < 2k3С). Отсюда можно сделать вывод, что для k3С < k3 < 2k3С отбор в сторону родственной кооперации будет слабее, чем для k3 < k3С, но при этом он все равно будет достаточно сильно действовать на популяцию. Результаты моделирования свидетельствуют в пользу этого предположения. На рис. 4а видно, что в отличие от первого интервала k3 < k3С, на котором численность различающих родственников агентов следует за численностью агрессивных агентов (т.е. практически все они вороны), на втором интервале k3С < k3 < 2k3С наблюдается сильный разброс значений частоты родственных агентов от эксперимента к эксперименту (см. также точки для стратегии ворона на рис. 4б).
Рис. 4. Значения усредненной частоты агрессивных[2] (кружки) и различающих родственников[3] (треугольники) агентов в популяции (а) и значения усредненной частоты стратегий голубей (крестики), воронов (кружки) и скворцов (треугольники) (б) для различных значений эффективности потребления ресурса k3. Частоты усреднялись по времени за период пребывания популяции в установившемся режиме. Линии проведены через средние значения. При k3 > 2k3С на эволюцию в популяции влияет несколько разнонаправленных факторов. За счет возможности кооперативной обороны скворцов, равновесие с высокой средней внутренней энергией по популяции, обычное для k3С < k3 < 2k3С, становится неустойчивым. Наличие скворцов создает отбор в сторону понижения энергии агента, такое понижение происходит за счет появления стратегий со все большей скоростью размножения. При этом если средний внутренний ресурс для агентов некоторой стратегии опускается ниже величины rs < k6, нападение на агентов этой стратегии становится не выгодно, так как в соответствии с определением взаимодействия между хищником и жертвой (см. таблицу 1), изменение ресурса хищника в этой ситуации становится отрицательным. Это делает возможным существование в популяции значительной доли голубей (рис. 4б). С другой стороны, конкуренция между голубями действует, как фактор отбора в сторону увеличения внутреннего ресурса, т.к. агент, имеющий больший запас энергии, имеет большее время на поиск ресурса. Все эти факторы приводят к сложной коэволюции стратегий голубей, воронов и скворцов. Высокая конкуренция как внутри, так и между стратегиями ведет к отбору кооперирующихся агентов. Результаты экспериментов показывают, что доля кооперирующихся агентов высока для всех реализаций (рис. 4а). При этом в отличие от первого интервала значений эффективности потребления ресурса k3 < k3С при k3 > 2k3С практически все голуби различают родственников (разность значений для агрессивных и кооперирующихся агентов на рис. 4а примерно равна численности голубей на рис. 4б). Детальное исследование динамики стратегий для высоких значений потребления ресурса представляет отдельный интерес и может выступать в качестве предмета дальнейших исследований. Третья глава посвящена моделированию социальной эволюции в примитивных сообществах охотников-собирателей и ранних земледельцев. Одним из основополагающих вопросов для теорий социальной эволюции можно считать вопрос об уровне кооперации, и ее механизмах в обществах с наиболее примитивной социальной организацией. Ответ на этот вопрос позволяет получить точку отсчета, с которой начинается социальная эволюция человеческих сообществ. Гипотеза, защищаемая в данной работе, состоит в том, что главным механизмом кооперации, влияющим на основные параметры примитивных социальных систем, является кооперация общин охотников-собирателей или ранних земледельцев на основе общности происхождения (родовых отношений). Для проверки гипотезы, предсказания, основанные на результатах вычислительных экспериментов с моделью, изложенной в предыдущей главе, проверяются на соответствие современным данным о примитивных обществах. Будем считать, что агенту из модели, изложенной в предыдущей главе, будет соответствовать община охотников собирателей или ранних земледельцев. Такая община, обычно состоящая из нескольких простых семей или одной расширенной семьи (семья с близкими родственниками), в первобытном обществе выступает в качестве отдельной автономной единицы. Члены общины селятся вместе, при перемещении с места на место и при взаимодействии с членами других общин, община выступает в качестве целостного субъекта. Людскому ресурсу, т.е. численности общины, будем сопоставлять значение внутреннего ресурса агента модели. При моделировании в качестве параметра, изменяющегося в серии экспериментов, выступала эффективность потребления ресурса, локализованного в клетке, одним агентом. Будем считать этот параметр аналогом количества ресурса (калорий), который может собрать община в окрестности стоянки, не перемещая ее. В качестве базового варианта был выбран третий вариант модели (см. табл. 2). Для оценки адекватности описания моделью реальных сообществ использовались две характеристики: 1. Частота в популяции агрессивных агентов. Агрессивными будем называть таких агентов, которые совершают действие a2 («нападать») хотя бы в одной из ситуаций, приведенных в таблице 3. Эта величина отражает количество агентов потенциально готовых совершать агрессивные действия. 2. Частота совершения действия f6 («отнять ресурс у соседа»). Дает оценку количества совершаемых агрессивных действий в популяции. Зависимость этих характеристик от величины ресурсной базы приведены на рисунках 5а и 5б соответственно. Эти зависимости демонстрируют, что поведение модели четко различается в трех интервалах: 1. k3 < k3С – скудная ресурсная база; 2. k3C < k3 < 2k3C – удовлетворительная ресурсная база; 3. k3 > 2k3C – хорошая ресурсная база. Отталкиваясь от результатов моделирования, можно сделать следующие предсказания о зависимости агрессивности в примитивных сообществах от экологических условий: 1. В условиях скудной ресурсной базы агрессивность агентов и количество совершаемых агрессивных актов увеличиваются по мере улучшения экологических условий. 2. При удовлетворительном обеспечении ресурсами частота агрессивных агентов в популяции максимальна, а количество агрессивных действий близко к нулю. При этом обе характеристики не зависят от количества ресурсов. 3. При хорошей ресурсной базе по мере ее увеличения, как частота агрессивных агентов, так и количество агрессивных действий растут. Согласно представлениям современной антропологии, подкрепленным богатым этнографическим материалом, улучшение ресурсной базы в большинстве случаев должно вести к повышению агрессивности в архаических обществах. Результаты моделирования находятся в соответствии с этой гипотезой в условиях скудной и хорошей ресурсной обеспеченности, но при промежуточных значениях параметра k3 модель обладает «аномальным» поведением. Таким образом, результаты моделирования «предсказывают», что, несмотря на низкий уровень межобщинной агрессии при удовлетворительной ресурсной базе, агенты должны обладать развитыми возможностями совершения агрессивных действий. Оказывается, что это действительно имеет место у охотников собирателей Калахари. Охотники племени !кунг всегда носят с собой отравленные стрелы, убить человека при помощи которых для охотника не представляет особого труда. Более того, существуют нормы, оправдывающие убийство посягнувшего на чужую собственность (улей с медом) [11]. Предсказание модели, подтверждается и наблюдениями аборигенов западной пустыни Австралии, которые живут в экологических условиях аналогичных условиям !кунг, и также демонстрируют низкий уровень агрессии. При этом у аборигенов присутствует институализация агрессии, проявляющаяся в виде создания отрядов тайных мстителей и ритуальных поединков [11].
Рис. 5. Значения усредненной частоты агрессивных агентов (а) и действия f6 (б) в популяции для различных значений эффективности потребления ресурса k3 (частота усреднялась по времени за период пребывания популяции в установившемся режиме, линии проведена через средние значения). Таким образом, результаты моделирования для первых двух интервалов обеспеченности ресурсом согласуются с данными наблюдений. Для того чтобы оценить адекватность модели при хорошей ресурсной базе, сравним ее с имеющимися представлениями и моделями конфликтности у ранних догосударственных земледельцев. Наиболее распространенная гипотеза о природе агрессивности у доиндустриальных обществ, основывается на мальтузианском предположении, заключающемся в том, что причинной повышения агрессии в обществе служит рост плотности населения. Следовательно, для реальных сообществ высокий уровень межобщинной агрессии должен положительно коррелировать с плотностью населения. Исходя из похожей посылки, Турчин и Коротаев предложили модель внутренней конфликтности для обществ, у которых отсутствует государственность [12]. Эта модель предсказывает наличие слабо отрицательной корреляции агрессии с плотностью населения. Расчет рассматриваемой корреляции для представленной в работе модели дает значения лежащие в интервале (-0.3;-0.6). Для того чтобы
протестировать представленные модели посмотрим, как плотность популяции коррелирует
с частотой конфликтов для примитивных обществ. Расчет корреляции между
переменной отражающей частоту межобщинных конфликтов и плотностью населения для
обществ, представленных в этнографической базе данных [13], дает результат Как показывают этнографические исследования, на межобщинный уровень агрессии влияет не только объем ресурсов, но и предсказуемость их доступности. Для догосударственных обществ корреляция между частотами стихийных бедствий (или неурожаев) и вооруженных столкновений составляет r = 0,631 (p = 0,046) [15]. Следовательно, увеличение вероятности появления ресурса в модели должно приводить к уменьшению показателей характеризующих агрессивность. Эксперименты с модифицированной моделью (вероятность увеличена в 10 раз) показывают, что агрессивность в модели падает в соответствии с наблюдаемой зависимостью, так при эффективности потребления ресурса k3 = 1500 обе характеристики агрессивности (см. выше) уменьшаются примерно в 3 раза. Четвертая глава содержит изложение модели эволюции целенаправленного поведения и результатов вычислительных экспериментов с ней. Целенаправленность одно из важнейших свойств поведения животных. В работе представлена модель эволюционного возникновения целенаправленного поведения основанного на мотивациях. Результаты компьютерных экспериментов с моделью показали, что в процессе эволюции в популяции агентов возникают устойчивые стратегии поведения, в которых информация о внутреннем ресурсе агента выступает в роли мотивации при выборе действий. Продемонстрировано, что мотивации играют важную роль в исследованных эволюционных процессах. А именно, если сравнить популяцию агентов с мотивациями, с популяцией агентов без мотиваций, то, как показывают компьютерные эксперименты, эволюционный процесс приводит к тому, что первая популяция (с мотивациями) имеет значительные эволюционные преимущества по сравнению со второй (без мотиваций). Также показано, что в процессе исследованных эволюционных процессов возникает естественная иерархическая структура целей и подцелей. Модели, предложенные в главе, являются отправной точкой в программе по построению серии моделей когнитивной эволюции [7]. Основные результаты диссертации1. Предложен новый способ формального описания моделей искусственной жизни, базирующийся на работах по теории автоматов. В рамках этого формализма на основе теории динамических систем были разработаны методы исследований данного класса компьютерных моделей. 3. Построена и исследована модель эволюции поведенческих стратегий в обществах, не имеющих сложной социальной организации. Показано, что изменение уровня предрасположенности к агрессии и частоты межобщинных конфликтов при изменении обеспеченности жизненными ресурсами находятся в согласии с данными об архаичных обществах. Проведено сравнение предсказаний модели о связи между частотой конфликтов и плотностью населения с предсказаниями альтернативных моделей и этнографическими данными, которое продемонстрировало, что предложенная в диссертации модель наиболее адекватна. 4. Построена и исследована модель эволюционного возникновения целенаправленного поведения, обусловленного мотивациями. Продемонстрировано, что мотивации играют важную роль в формировании адаптивного поведения. По теме диссертации опубликованы следующие работы:1. Burtsev M.S. Tracking the Trajectories of Evolution // Artificial Life 10(4), 2004. стр. 397-411. 2. Бурцев М.С., Гусарев Р.В., Редько В.Г. Исследование механизмов целенаправленного адаптивного управления // Известия Академии Наук. Теория и системы управления. т. 6. 2002. стр. 55-62. 3. Бурцев М.С., Гусарев Р.В., Редько В.Г. Модель эволюционного возникновения целенаправленного адаптивного поведения 1. Случай двух потребностей // Препринт ИПМ РАН № 43. М., 2000. 24 стр. 4. Бурцев М.С. Модель эволюционного возникновения целенаправленного адаптивного поведения. 2. Исследование развития иерархии целей // Препринт ИПМ РАН № 69. М., 2002. 15 стр. 5. Burtsev M.S. Measuring the Dynamics of Artificial Evolution // Lecture Notes in Computer Science, 2801 / 2003. Advances in Artificial Life. 2003. стр. 580-587. 6. Burtsev M.S., Red'ko V.G., Gusarev R.V. Model of Evolutionary Emergence of Purposeful Adaptive Behavior. The Role of Motivation // Advances in Artificial Life: 6th European Conference, ECAL 2001, Prague, Czech Republic, Proceedings. Springer-Verlag Heidelberg. 2001. 7. Red'ko V.G., Prokhorov D.V., Burtsev M.S. Theory of Functional Systems, Adaptive Critics and Neural Networks // Proceedings of IJCNN 2004. стр. 1787-1792. 8. Burtsev M.S. Artificial Life Meets Anthropology: A Case of Aggression in Primitive Societies // Proceedings of ECAL 2005. 9. Редько В.Г., Бурцев М.С., Гусарев Р.В. Проблема моделирования эволюционного происхождения интеллекта // Сб. труд. конф. "Биосфера и человечество". Обнинск, МРНЦ РАН. 2000. стр. 123-129. 10. Бурцев М.С., Гусарев Р.В., Редько В.Г. Эволюционное возникновение целенаправленного поведения в модели искуственно жизни // Математика. Компьютер. Образование. Вып.8/ Тезисы. Пущино, 2001. 11. Бурцев М.С., Гусарев Р.В., Редько В.Г. Нейросетевая модель эволюционного возникновения целенаправленного адаптивного поведения // III Всероссийская научно-техническая конференция "Нейроинформатика-2001". М., МИФИ. 2001. стр. 153-160. 12. Бурцев М.С. Формирование иерархии целей в модели искусственной эволюции // IV Всероссийская научно-техническая конференция "Нейроинформатика-2002". М., МИФИ. 2002. 13. Бурцев М.С. Эволюционно-кибернетический подход к моделированию адаптивного поведения // 8-ая национальная конференция по искусственному интеллекту с международным участием. М., Физматлит. 2002. стр. 790-798. 14. Анохин К.В., Бурцев М.С., Зарайская И.Ю., Лукашев А.О., Редько В.Г. Проект "Мозг Анимата": разработка модели адаптивного поведения на основе теории функциональных систем // 8-ая национальная конференция по искусственному интеллекту с международным участием. М., Физматлит. 2002. стр. 781-789. 15. Бурцев М.С., Редько В.Г. Влияние агрессии на эволюцию в многоагентной системе // Сб. трудов 9-ой Международной конференции "Проблемы управления безопасностью сложных систем". М., ИПУ. 2002. 16. Бурцев М.С. Исследование эволюции адаптивного поведения в модели искусственной жизни // Труды Международных конференций "Искусственные интеллектуальные системы" (IEEE AIS'02) и "Интеллектуальные САПР" (CAD-2002). М., Физматлит. 2002. стр. 514-520. 17. Редько В.Г., Крыжановский М.В., Бурцев М.С., Анохин К.В. Теория функциональных систем как концептуальная основа исследований интеллектуального адаптивного поведения // Интеллектуальные и многопроцессорные системы – 2003. Таганрог, Издательство ТРТУ, 2003. Т.1. стр. 60-63. 18. Бурцев М.С. Что такое Искусственная жизнь? // Труды XLVI научной конференции МФТИ "Современные проблемы фундаментальных и прикладных наук". Часть I. Радиотехника и кибернетика. Москва – Долгопрудный, МФТИ. 2003. стр. 17-19. 19. Мосалов О.П., Редько В.Г., Бурцев М.С., Митин Н.А. Модель эволюционирующей многоагентной Интернет-системы // Интегрированные модели и мягкие вычисления в искусственном интеллекте. Сборник трудов II-го Международного научно-практического семинара. М., Физматлит. 2003. стр. 217-221. 20. Мосалов О.П., Бурцев М.С., Митин Н.А., Редько В.Г. Модель многоагентной Интернет-системы, предназначенной для предсказания временных рядов. // V Всероссийская научно-техническая конференция "Нейроинформатика-2003". М., МИФИ. 2003. стр. 177-183. 21. Редько В.Г., Мосалов О.П., Митин Н.А., Бурцев М.С. Многоагентные Интернет-системы – исследование процессов обучения и самоорганизации // Сб. трудов Первой Всероссийской научной конференции «Методы и средства обработки информации». М., МГУ. 2003. стр. 280-285. 22. Burtsev M.S. Evolution of strategies in the Artificial Life model: an attempt of historical interpretation // тезисы Международной конференции "Математическое моделирование социальной и экономической динамики". М.: МГСУ, 2004. 23. Burtsev M.S. Does evolution of kinship organization lead to pacification? // тезисы 3-ей Международной конференции "Иерархия и власть в истории цивилизаций" М.: Иаф РАН, 2004. 24. Бурцев М.С. Классическая и эволюционная причинность в моделях обучения // 9-ая национальная конференция по искусственному интеллекту с международным участием. Труды конференции. М., Физматлит. 2004. 25. Бурцев М.С. Эволюция кооперации в многоагентной системе // VII Всероссийская научно-техническая конференция "Нейроинформатика-2005". М., МИФИ. 2005. 26. Бурцев М.С. Метафизика Дарвина против метафизики Ньютона: два подхода к причинности в обучении // Всероссийская междисциплинарная конференция «Философия искусственного интеллекта». М., МИЭМ. 2005. Список литературы:1. Нейман Дж. Теория самовоспроизводящихся автоматов. – М.: Мир, 1971. 2. Цетлин М.Л. Исследования по теории автоматов и моделированию биологических систем. – М.: Наука, 1969. 3. Кулагина О.С., Ляпунов А.А. К вопросу о моделировании эволюционного процесса // сб. "Проблемы кибернетики", вып. 16. M.: Наука, 1966. 4. Evolution, Games and Learning: Models for Adaptation in Machines and Nature // Physica D 22D (1-3), 1986. 5. Новое в синергетике. Взгляд в третье тысячелетие. Сборник статей под ред. Г.Г.Малинецкого, С.П.Курдюмова. – М.: Наука, 2002. 6. Куракин П.В., Малинецкий Г.Г. На пороге "субъективной" синергетики (синергетика II) // Синергетика. Труды семинара. Т.3, М., изд-во МГУ, 2000. С.242-250 7. Редько В.Г. Эволюционная кибернетика. – М.: Наука, 2001. 8. Holland J.H. Adaptation in natural and artificial systems: an introductory analysis with applications to biology, control, and artificial intelligence. – Cambridge, Massachusetts: MIT Press, 1992. 9. Hamilton W.D. The evolution of altruistic behavior // Am. Nat. 97, 1963. C. 354-356. 10. Maynard Smith J. Evolution and the Theory of Games. Cambridge: Cambridge University Press, 1982. 11. Казанков А.А. Агрессия в архаических обществах. – М .: Иаф РАН, 2002. 12. Turchin P., Korotayev A. Population Dynamics and Internal Warfare: a Reconsideration // American Anthropologist (в печати). 13. Standard Cross-Cultural Sample Variables (Edited by William T. Divale, Daria Khaltourina and Andrey Korotayev) // World Cultures 13, 2002. (файлы STDS01-88.SAV) 14. Nolan P.D. Toward an Ecological-Evolutionary Theory of the Incidence of Warfare in Preindustrial Societies // Sociological Theory V.21, N1, p.18-30, 2003. 15. Ember C.R., Ember M. Resource Unpredictability, Mistrust, and War: A Cross-Cultural Study // Journal of Conflict Resolution V.36, p.242-262, 1992. [1] Названия стратегий предложены П.В. Турчиным. [2] Агрессивными считались агенты, которые совершали действие a2 ("нападать") хотя бы в одной из ситуаций из таблицы 3. [3] Считалось, что агент отличает родственников, если хотя бы в одной из ситуаций из таблицы 3 действие в отношении родственника отличалось от действия в отношении не родственника. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
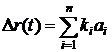
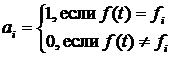 . (5)
. (5)