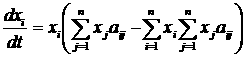Пасквиль на эволюцию[1]Бурцев М.С., к.ф.-м.н., Малинецкий Г.Г., д.ф.-м.н. Институт прикладной математики им. М.В.Келдыша РАН "С. Лем СУММА ТЕХНОЛОГИИ ГЛАВА ВОСЬМАЯ ПАСКВИЛЬ НА ЭВОЛЮЦИЮ КОНСТРУКЦИИ, ОСНОВАННЫЕ НА ОШИБКАХ"
Искусственная жизнь, как метод компьютерного моделирования нелинейных процессов«Искусственная жизнь» – молодое междисциплинарное направление исследований. Предметом исследования искусственной жизни является эволюция[2] сложных систем, а основным методом – построение компьютерных моделей. Модели искусственной жизни находят свое применение в теоретических областях исследований возникновения жизни, биологической эволюции, коллективного поведения, социальной эволюции. Чтобы понять место искусственной жизни среди других методов, рассмотрим основные этапы развития математического моделирования. Возникновение математического моделирования в его современном виде связано с изобретением в XVI веке дифференциального исчисления Ньютоном. На первом этапе его развитие было связано с методами интегрирования систем дифференциальных уравнений. При этом считалось, что любую систему, описывающую динамику, можно свести к интегрируемой. Это позволило решить огромное число задач механики, для которых имеется возможность построить интегрируемую систему, и заложить фундамент современной физики. Следующий этап развития математического моделирования связан с проблемой неинтегрируемости системы трех взаимодействующих тел. Многочисленные попытки свести эту систему к интегрируемой оказались бесплодными и были прекращены Брунсом и Пуанкаре в конце XIX века, доказавшими невозможность подобного преобразования. Открытие целого класса неинтегрируемых систем привело Пуанкаре к созданию качественной теории дифференциальных уравнений (теории динамических систем) для их исследования. Так как для неинтегрируемых систем невозможно получить аналитического решения, то объектом исследования теории динамических систем становятся качественные свойства решений, а не сами решения. Основным понятием новой теории стало понятие фазового пространства. Фазовое пространство состоит из точек, представляющих различные состояния системы, а его преобразования определяются динамикой системы. Фазовое пространство можно разбивать на области в зависимости от того, куда, в конце концов, попадают траектории начинающиеся в них. Множества в фазовом пространстве, «притягивающие» траектории, получили название аттракторов. Качественная теория дифференциальных уравнений открыла дорогу широкому использованию нелинейных систем в физике. Исследования диссипативных процессов теплопроводности, диффузии, вязкости и т.д. в открытых нелинейных системах привели к возникновению физики неравновесных систем [1] и синергетики [2]. Оказалось, что многие процессы в природе приводят к образованию диссипативных структур – стационарных распределений значений переменных, обладающих устойчивостью к возмущениям. Движение системы, ведущее к подобному стационарному распределению, получило название самоорганизации. На этом этапе в центре внимания нелинейной динамики оказались бифуркации – изменение числа и устойчивости решений (в частности, конфигураций диссипативных структур) при вариации параметров системы. Синергетика дала возможность моделирования образования пространственных структур, колебательных химических реакций, неравновесных систем и т.д. Следующий период истории математического моделирования нелинейных систем связан с динамическим хаосом. Выяснилось, что простые уравнения могут порождать сложную динамику, важной характеристикой которой оказалась степень ее предсказуемости – горизонт прогноза [3]. Было показано, что существуют простые динамические системы, поведение которых на больших интервалах времени невозможно предсказать. Примером хаотического поведения является турбулентность, принципиально ограничивающая возможности прогнозирования погоды. Исследования динамического хаоса нашли практическое применение в таких областях, как создание генераторов шума, шифрование и компрессия данных. Сегодня в математическом моделировании нелинейных процессов происходит становление новой парадигмы – парадигмы сложности. Ее появление вызвано нарастающей потребностью в создании моделей «сложных» систем – биологических, социальных, экономических, политических, исторических. Эта потребность не была полностью удовлетворена уже имеющимися инструментами моделирования, что повлекло за собой создание широкого междисциплинарного направления исследований, призванного синтезировать подходы нелинейной динамики, теории динамического хаоса и вычислительного эксперимента. Характерными свойствами объектов моделирования в новой парадигме становятся: необратимость эволюции, редкие катастрофические события, высокая гетерогенность и адаптивность взаимодействующих элементов системы. Результаты исследований, проведенных в рамках парадигмы сложности, уже вылились в ряд важных приложений, таких как искусственные нейронные сети в области управления и предсказания, генетические алгоритмы в области оптимизации. Одно из основных теоретических течений в парадигме сложности – это искусственная жизнь. Исследования в области искусственной жизни являются продолжением пионерских работ по теории самовоспроизводящихся автоматов [4], игр автоматов [5] и компьютерного моделирования эволюции [6]. С момента своего возникновения, искусственная жизнь тесно связана с синергетикой и нелинейной динамикой, не случайно первый сборник статей в этой области вышел в журнале Phisyca D «Nonlinear Phenomena» [7]. И сегодня синергетика, как междисциплинарный подход, и искусственная жизнь, как одно из направлений математического моделирования, широко используют представления о самоорганизации и динамических системах, и имеют общее направление движения [8,9]. Методы искусственной жизни активно используются в работах по моделированию эволюции когнитивных способностей животных [10], в областях эволюционной роботики и эволюционных вычислений [11]. Модели искусственной жизни относятся к классу многоагентных (“multi-agent”, “agent-based”) компьютерных моделей. Принципиальным их отличием от других многоагентных моделей является обязательное наличие эволюции свойств агентов через наследование со случайными мутациями. Таким образом, метод искусственной жизни изначально является стохастическим, и его можно рассматривать, как подмножество методов Монте-Карло. Потребность в моделях искусственной жизни возникает, когда из-за сложности и разнообразия поведения отдельных индивидуумов (например, в зависимости от локального окружения) трудно или невозможно получить систему уравнений, описывающих динамику популяции. При этом сами результаты компьютерных экспериментов могут помочь в поиске приемлемых аналитических моделей. Кроме того, многоагентное моделирование может быть использовано в качестве альтернативы или для верификации результатов численного решения системы уравнений. Современная наука остро нуждается в новых инструментах для теоретических и прикладных исследований сложных систем. Искусственная жизнь, базирующаяся на эволюционной идеологии и синтезе методов нелинейной динамики, синергетики и компьютерного моделирования, претендует на роль одного из таких инструментов. Искусственная жизнь, как метод моделирования эволюцииЭволюционная теория оказала и продолжает оказывать значительное влияние не только на биологию, но и на другие области науки. Больцман утверждал, что при разработке термодинамики он пытался построить теорию аналогичную дарвиновской, но только в физике. В философской теории познания дарвинизм привел к эволюционной эпистемологии [12,13], а в экономике – к эволюционной экономике [14,15]. Классическая теория игр [16,17], ориентированная на поиск статической оптимальной стратегии, сначала была применена в теоретической биологии для моделирования эволюции в популяции [18-20], а затем полученные результаты были использованы и для развития самой теории игр, но уже в направлении исследования динамики стратегий. Эта ветвь теории игр называется эволюционная теория игр [21] и посвящена динамической устойчивостью стратегий. Эволюционная теория нашла свое применение и в исследованиях человеческого общества. В современных социальных науках сформировалось целое направление, прошедшее путь от «Агрессии» Лоренца [22] и «Социобиологии» Уилсона [23] до современных теорий коэволюции генов и культуры [24-26]. Математизация эволюционной теории началась в области генетики популяций в начале 20-го века [27-29]. Вопрос, на который должна была ответить теория, можно сформулировать так: Каковы будут изменения в генетическом составе популяции при данной стратегии размножения и заданных постоянных значениях приспособленности для каждого гена? При такой постановке вопроса, очевидный ответ состоял в том, что частота гена в популяции увеличивается со скоростью, пропорциональной приспособленности. Вероятно, неочевидность способа экспериментального определения приспособленности в совокупности с простотой модельных представлений привели к тому, что произошла подмена причины и следствия. Вместо дарвиновской логической цепочки – приспособленность определяет скорость воспроизводства вида – распространилось определение приспособленности, как скорости воспроизводства вида, – т.е. скорость воспроизводства вида есть приспособленность [30]. Положение дел начало меняться при заимствовании метода теории игр и создании эволюционной теории игр. В эволюционной теории игр воспроизводство вида определяется значением выигрыша некоторого вида при взаимодействии с другими видами в популяции. Таким образом, проблему эволюционной теории игр можно сформулировать следующим образом: Какова будет динамика частот стратегий в популяции при заданных результатах взаимоотношений между стратегиями (матрице выигрышей)? Понятие приспособленности при таком подходе утрачивает смысл, вкладываемый в него популяционной генетикой. Рассмотрим динамику в некоторой популяции: пусть после некоторого переходного периода в популяции устанавливается эволюционно устойчивое распределение по стратегиям. Можно считать, что именно в этом состоянии все виды наиболее приспособлены друг к другу. Однако такое состояние совсем не обязательно гарантирует максимизацию среднего по популяции выигрыша. И как не парадоксально, но стратегии, обеспечивающие разный выигрыш, имеют (в смысле классической популяционной генетики) одинаковые относительные и абсолютные приспособленности (равные нулю и единице соответственно). Основное уравнение эволюционной теории игр – уравнение репликации:
где xi – частота i-ой стратегии, aij – выигрыш i-ой стратегии при взаимодействии с j-ой стратегией. Это уравнение эквивалентно уравнению Лотки-Вольтерры [31,32], которое используется для моделирования экологических взаимодействий. Ни уравнение репликации, ни уравнение Лотки-Вольтерры не требует введения понятия приспособленности. В первом случае эволюция определяется матрицей выигрышей, а во втором – скоростью воспроизведения вида и матрицей его взаимодействий с другими видами. Это наводит на мысль о том, что понятие приспособленности не является необходимым при моделировании эволюционной динамики. Следовательно, исследование модели сводится к анализу возможных путей ее эволюционного движения от одного относительно устойчивого динамического режима к другому [33,34]. Несмотря на это, обычной практикой остается использование понятия приспособленности при рассмотрении любых эволюционных моделей, включая эволюционную теорию игр и уравнение Лотки-Вольтерры [35]. В развитии современной эволюционной теории можно проследить переход от вопросов, связанных с эволюцией популяции на заданном ландшафте приспособленности[3], к вопросам о том, как зависит ландшафт приспособленности от взаимодействий между различными особями и внешних условий. Это нашло отражение в постепенном включении в эволюционную теорию сначала моделей родственного отбора[4] [36-38], группового отбора[5] [38-40] и неслучайных взаимодействий[6] [41-43], а затем пространственной структуры популяции [44,45]. Появление сложных эволюционных моделей привело к широкому использованию вычислительных экспериментов и созданию отдельного направления исследований, которое получило название искусственная жизнь [7,46-47]. В искусственной жизни эволюционирующая система задается исходя из некоторой «элементарной аксиоматики», определяющей физические свойства мира, «заселенного» агентами, и свойства самих агентов. Агенты способны размножаться и гибнуть. В течение жизни поведение отдельного агента определяется алгоритмом, унаследованным от родителей. При наследовании алгоритм передается потомку не точно, а с ошибками, играющими роль аналогичную биологическим мутациям. Важным отличием искусственной жизни от других подходов к моделированию эволюции является то, что это направление было изначально ориентировано на компьютерное моделирование как основной инструмент исследования [7]. Это значительно облегчило построение моделей с богатым спектром сценариев эволюционного развития. Основные отличия искусственной жизни от основных методов эволюционной биологии – теории игр и популяционной генетики заключаются в следующем: · моделирование как генотипа, так и фенотипа. В популяционной генетике фенотип вынесен за скобки, исследуются только изменения генных частот. Напротив, в эволюционной теории игр генотип отсутствует, а моделируется динамика фенотипических признаков – стратегий. В искусственной жизни агент имеет и генотип, определяющий фенотипические признаки, и фенотип, от которого зависит выживание агента. Наличие, как генотипа, так и фенотипа позволяет исследовать, как отображение из первого во второй влияет на эволюцию. · разнообразие генетических операторов. Так как каждый из агентов в искусственной жизни обладает достаточно сложным набором генов, то становится возможным изучение влияния различных генетических операторов (мутаций, кроссовера и инверсий) и типов размножения (бесполое и половое) на эволюцию. · широкий спектр игр – ситуаций. В эволюционной теории игр обычно рассматриваются игры, в которых каждый из игроков может сделать один из двух ходов, например, кооперироваться или не кооперироваться. В искусственной жизни репертуар действий агентов и изменений во внешней среде гораздо разнообразней, что создает большее число ситуаций, в которых агент вынужден выбирать то или иное действие. · разнообразие стратегий поведения. Значительное число ситуаций, в которых может находиться агент, и действий, которые он может совершать, вынуждает использовать «интеллектуальные» алгоритмы для управления поведением агента, такие как искусственные нейронные сети или нечеткую логику. К сожалению, богатство динамики моделей искусственной жизни значительно осложняет анализ и интерпретацию результатов численных экспериментов. Слабая проработанность этого вопроса является основным недостатком искусственной жизни сегодня. Для преодоления этой «болезни роста» необходимо накопление более широкого опыта применения данного класса моделей в различных областях науки. Применение искусственной жизни позволяет подвергнуть теоретические эволюционные концепции более жесткому тесту. Так, в исследованиях эволюции кооперации, базирующихся на наиболее распространенном сегодня подходе теории игр, репертуар возможных взаимодействий между агентами очень беден, обычно агент может выбрать одно из двух действий – кооперироваться или не кооперироваться с текущим партнером. Искусственная жизнь позволяет смоделировать эволюцию популяции с гораздо более широкими и естественными возможностями для взаимодействия агентов, что дает гораздо более широкий спектр стратегий поведения агентов. Исследование такой модели кооперации [48,49] позволило увидеть не только конкуренцию малого количества заранее заданных кооперативных и эгоистических стратегий поведения (единицы стратегий), но сам сценарий возникновения кооперативных стратегий (всего возможно более 101000 различных стратегий) и то, какие из кооперативных стратегий будут отобраны эволюцией. Несмотря на все сложности с анализом моделей искусственной жизни сегодня они популярны не только в биологии, но и в социальных науках [50-52]. Среди других биологических систем человеческие сообщества обладают наибольшей гетерогенностью агентов и сложностью их индивидуального поведения. Как уже говорилось выше, это затрудняет построение моделей традиционными методами. Однако, использование искусственной жизни для исследования социальных систем, также как и любого другого подхода в данной области, не позволяет избежать трудностей с корректностью построения модели и интерпретацией результатов. Для того чтобы исследовать возможность использования искусственной жизни для моделирования реальных социальных систем, была предпринята попытка построить модель для предсказания основных характеристик и динамики эволюции в примитивных эгалитарных сообществах охотников-собирателей и ранних земледельцев [53,54]. Проверка предсказаний, следующих из результатов моделирования, на этнографических данных показала их адекватность. Как и при исследовании других моделей, основной задачей при работе с моделями искусственной жизни является изучение зависимости решения (результатов моделирования) от параметров модели и начальных условий. При этом получение данных о поведении многоагентной системы затруднено необходимостью привлечения значительных вычислительных ресурсов. Это один из недостатков моделей искусственной жизни. К тому же для подобных моделей не всегда сразу очевидна связь начальных допущений с результатом, так как динамика эволюционирующей системы определяется процессами самоорганизации. Для выявления связи начальных допущений модели с получаемым поведением требуются дополнительные вычислительные эксперименты, что приводит к увеличению времени, затрачиваемого на исследования. Искусственная жизнь – современный метод исследования процессов эволюции сложных систем, появившийся благодаря развитию вычислительных возможностей человека. Пусть иногда модели искусственной жизни кажутся со стороны всего лишь «пасквилем на эволюцию», они помогают нам лучше понять эволюционные процессы, и, как ни странно, сами являются не только плодом, но и инструментом дальнейшего эволюционного развития. Литература1. Николис Г., Пригожин И. Самоорганизация в неравновесных системах. – М.: Мир, 1979. 2. Хакен Г. Синергетика. – М.: Мир, 1980. 3. Пределы предсказуемости / Сб. статей под ред. Ю.А. Кравцова. М.: Центрком, 1997. 4. Нейман Дж. Теория самовоспроизводящихся автоматов. – М.: Мир, 1971. 6. Кулагина О.С., Ляпунов А.А. К вопросу о моделировании эволюционного процесса // сб. «Проблемы кибернетики», вып. 16. M.: Наука, 1966. 9. Куракин П.В., Малинецкий Г.Г. На пороге «субъективной» синергетики (синергетика II). Синергетика. Труды семинара. Т.3, С.242-250. М.: изд-во МГУ, 2000. 10. Редько В.Г. Эволюционная кибернетика. – М.: Наука, 2001. 11. Holland J.H. Adaptation in natural and artificial systems: an introductory analysis with applications to biology, control, and artificial intelligence. – Cambridge, Massachusetts: MIT Press, 1992. 12. Эволюционная эпистемология и логика социальных наук: Карл Поппер и его критики. Под ред. Садовского В.Н. – M.: Эдиториал УРСС, 2000. 13. Эволюция, культура, познание. - М., 1996. 14. Nelson R.R., Winter S.G. An Evolutionary Theory of Economic Change. – London: Belknap Press, 1982. 15. The Economy as an Evolving Complex System // Anderson P.W., Arrow K.J., Pines, D. (eds.), Addison-Wesley, Reading, Mass., 1988. 16. von Neumann J., Morgenstern O. Theory of Games and Economic Behaviour. – Princeton: Princeton University Press, 1947. 17. Nash J. Non-cooperative games // Ann. Math. V.54, p.287-295, 1951. 18. Maynard Smith J. The theory of games and the evolution of animal conflicts // J. Theor. Biol. V.47, p.209-221, 1974. 19. Maynard Smith J. Evolution and the Theory of Games. – Cambridge: Cambridge University Press, 1982. 20. Hofbauer J., Sigmund K. Evolutionary Games and Population Dynamics. – Cambridge: Cambridge University Press, 1998. 21. Hofbauer J., Sigmund K. Evolutionary game dynamics // Bull. Am. Math. Soc. V.40, N4, p.479-519, 2003. 22. Lorenz K. On Aggression. – London: Methuen, 1966. 23. Wilson E.O. Sociobiology: The New Synthesis. – Cambridge, MA: Harvard University Press, 1975. 24. Boyd R., Richerson P.J. Culture and the evolutionary process. – Chicago: University of Chicago Press, 1985. 25. Sober E., Wilson D.S. Unto Others: The Evolution and Psychology of Unselfish Behavior. – Cambridge, MA: Harvard University Press, 1998. 26. Richerson P.J., Boyd R., Henrich J. The Cultural Evolution of Human Cooperation // The Genetic and Cultural Evolution of Cooperation, P. Hammerstein ed., p.357–388. Cambridge MA: MIT Press, 2003. 27. Fisher R.A. The genetical theory of natural selection. – Oxford: Clarendon Press, 1930. 28. Haldane, J. B. S. A mathematical theory of natural and artificial selection. Parts I-V // Transactions of the Cambridge Philosophical Society V.23, 1924-1927. 29. Wright S. Evolution in Mendelian populations // Genetics V.16, p.97-159, 1931. 30. Ariew A., Lewontin R. C. The Confusions of Fitness // Brit. J. Phil. Sci. V.55, p.347–363, 2004. 31. Lotka A. J. Undamped oscillations derived from the law of mass action // J. Am. Chem. Soc. V.42, p.1595–1599, 1920. 32. Volterra V. Variozioni e fluttuazioni del numero d’individui in specie animali conviventi // Mem. Accad. Naz. Lincei V.2, p.31–113, 1926. 34. Burtsev M.S. Tracking the Trajectories of evolution // Artificial Life V.10, p.397-411, 2004. 35. Page K.M., Nowak M.A. Unifying Evolutionary Dynamics // J. theor. Biol. V.219, p.93–98, 2002. 36. Hamilton W.D. The evolution of altruistic behavior // Am. Nat. V.97. p.354-356, 1963. 37. Hamilton W.D. The genetical evolution of social behavior, I and II // J. Theor. Biol. V.7, p.1-52, 1964. 38. Maynard Smith J. Kin selection and group selection // Nature V.201, p.1145-1147, 1964. 39. Wright S. Tempo and Mode in Evolution: A Critical Review // Ecology V.26, N.4, p.415-419, 1945. 40. Wilson D.S. A theory of group selection // Proc. Nat. Acad. Sci. USA. V.72, N1, p.143-146, 1975. 41. Trivers R. The evolution of reciprocal altruism // Q. Rev. Biol. V.46, p.35-57, 1971. 42. Wilson D.S., Dugatkin L.A. Group selection and assortative interactions // Amer. Natur. V.149, N2, p.336-351, 1997. 43. Eshel I., Cavalli-Sforza L.L. Assortment of encounters and evolution of cooperativeness // PNAS (Part 1: Biological Sciences) V.79, N4, p.1331-1335, 1982. 44. Nowak M.A. May R. M. Evolutionary games and spatial chaos // Nature V.359, p.826-829, 1992. 45. Brauchli K., Killingback T., Doebeli M. Evolution of cooperation in spatially structured populations // J. theor. Biol. V.200, p.405-417, 1999. 46. Artificial Life // Langton C.G. ed. – MA: The MIT Press, 1995. 47. Adami C. Introduction to Artificial Life. – NY: Springer Verlag, 1998. 48. Бурцев М.С. Искусственная жизнь как метод математического компьютерного моделирования процессов эволюции сложных систем // Информационные технологии и вычислительные системы, N 3, стр. 5 – 22, 2005. 49. Burtsev M.S., Turchin P.V. Evolution of cooperative strategies from first principles // Nature 440, 1041-1044, 2006. 51. Lansing J.S. “Artificial Societies” and the Social Sciences // Artificial Life V.8, p.279-292, 2002. 53. Бурцев М.С. Эволюция кооперации в компьютере и социуме // Новое в синергетике. Новая реальность, новые проблемы, новое поколение. Сборник статей. Ч. 2. Малинецкий Г.Г. (Ред.), стр. 40-56, 2006. 54. Burtsev M.S. Artificial Life Meets Anthropology: A Case of Aggression in Primitive Societies // In M. Capcarrere et al. (Eds.): ECAL 2005, LNAI 3630, pp. 655 – 664, 2005. [1] Работа поддержана грантом РФФИ №05-01-00852 [2] Необходимо отметить, что значение термина эволюция в искусственной жизни и в области математического моделирования различаются. Если в последнем случае под эволюцией подразумевается динамика исследуемой системы во времени, то в искусственной жизни это процесс «развития» системы, ее усложнения (или упрощения), имеющий принципиально стохастический характер. [3] Приспособленность организма можно представить в виде функции многих переменных, каждая из которых определяется состоянием некоторого гена. В пространстве всех возможных генотипов эта функция будет задавать поверхность, называемую в эволюционной теории ландшафтом приспособленности. Эволюцию популяции можно представить, как движение по ландшафту приспособленности от долин (низкая приспособленность) к пикам (высокая приспособленность). [4] Эволюционные механизмы, приводящие к возникновению кооперативного поведения между родственниками, называются родственным отбором. [5] Вызванное естественными факторами разбиение популяции на небольшие субпопуляции, ведет к значительному повышению генетической неоднородности и конкуренции между локальными группами, и, как следствие, к ускорению эволюции. Это явление получило название группового отбора. [6] В классической популяционной генетике предполагается, что взаимодействия между особями внутри популяции случайны, введение в теорию зависимости вероятности взаимодействия двух индивидов от их генов позволяет получить более реалистичные результаты. |